
By John Conway
When setting out to restore fossil animals, an artist is faced with a unique problem that is not often faced by scientists. Where a scientist can reasonably reach the conclusion that something is not—or cannot be— known, a palaeontological artist is forced to reach conclusions on things which are unknown, and perhaps always will be. An artist can't paint a pterosaur in "don't know" colours, or behaving in a "don't know" way. Restoring pterosaurs, therefore, not only requires research into what is known about pterosaur's anatomy, behaviour, and environment, but also a method for building up complete picture, even when there is little or no evidence to be had.
Restoring Anatomy
There are four main anatomical systems of interest to an artist: the skin and integument, the musculature, the pulmonary system, and the skeleton. All these systems had an impact on a pterosaur shape and appearance.

Skeleton :: Musculature :: Pulmonary :: External
The building up the four main structural systems ofthe pterosaur Anhanguera, layer by layer.
(Sources: Bennett 2003, Kellner & Tomida 2000, Hudson 1955, Ritchison 2007, Meers 2003, Unwin 2006, Wellnhofer 1991, Witmer 2003)

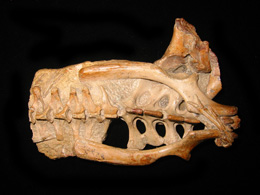
Pterosaur anatomy is reconstructed from fossils such as these: a beautifully articulated but flat Pterodactylus from Solnhofen (top), and a three-dimensionally preserved but partly disarticulated Anhanguera pelvis from the Santana formation, Brazil (bottom).
Restoring the anatomy of any pterosaur starts with the fossil remains. Most pterosaurs are only known from bones, but sometimes soft-tissue impressions present as well. To pin down the general proportions, it is necessary to draw a skeletal diagram. It's best to trace the bones directly from photographs or the original descriptions of the fossils.
Fossil material is usually incomplete and often crushed, so an attempt has to be made to fill in the gaps, and correct for the distortion caused by crushing. Missing material can often be filled with material that is known from related species, but care must be taken choose and appropriate relative (see origins and relationships), and to account for possible differences (such as the proportional changes related to size).
When drawing a skeletal diagram, it is essential that all the bones are articulated properly, and within their natural range of motion. This is not a trivial task, as many pterosaur fossils have poorly preserved joints, and distorted, crushed bones. Articulated specimens such as Pterodactylus (right, top) and uncrushed 3D specimens such as Anhanguera (right, bottom) can be especially helpful in this regard.
The musculature of pterosaurs can be inferred from muscle scars on the bones, and comparison with related living animals, such as crocodiles, birds, and lizards. This method is called phylogenetic bracketing. The most prominent muscle in pterosaurs would have been the pectoralis - the large muscle that ran from the upper arm to the sternum, and pulled the wing down. The other muscles of arm may have been quite prominent as well - such as the biceps, and the forearm muscles which bunched around the elbow, and controlled the flight finger (Bennett 2003, 2007).
Pterosaur skin is preserved in several specimens. Skin in known from the wing membranes, throat, head-crests and feet. On the wings the skin is stiffened by fibres known as actinofibrils, which were the consistency of coarse hair, and had an aerodynamic function. The skin of the throat and head crests reveals a slightly wrinkled non-scaley surface. Impressions from the feet indicate small pebbly non-overlapping scales were present on the heel.
Many fossils also preserve hair-like integument, which is often called "ptero-fuzz". Ptero-fuzz varied from coarse to fine, and seems to have been extensive in at least some pterosaurs. The fuzz may have been dry and truly "fuzzy" as in bats, or it may have been kept oily and slicked, as bird feathers. Certainly have an oily coat would have helped prevent fishing pterosaurs from getting water-logged if they came in contact with the water.
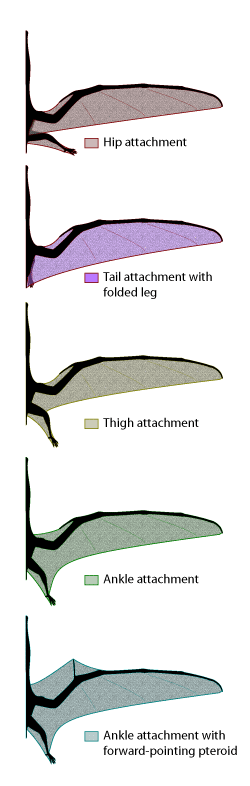
Five of the many various wing-membrane configurations that have been proposed for pterosaurs.
Historically, the extent and shape of pterosaur membranes has been controversial, and remains so to this day. Many wing-membrane impressions are known, but they are often frustratingly ambiguous as to the attachment to the body and hind leg. Most scientists prefer the ankle attachment scenario, which is clearly seen in some pterosaur fossils. Others maintain that there might have been a variety of attachments, and that having the hindlimb free of the main wing would have aerodynamic advantages for the large dynamic soarers (such as Anhanguera and Pteranodon).
Pterosaurs evolved a remarkable array of head-crests, from the keeled jaws and snout of Anhanguera, to the huge sail-like crest of Tapejara, and may more besides. Nearly all pterodactyloid pterosaurs had a head-crest of some description, and soft tissue preservation inticates that bony crests were often significantly extended with horn and skin. Many of these crests are believed to have been used for display, and therefore may have been brightly coloured or patterned.
Restoring Behaviour
The behaviour of pterosaurs is obviously highly speculative - very little can be known for certain, and much will never be known. In restoring pterosaurs, it is perhaps fair to say that anything that doesn't seem highly unlikely is valid speculation. The two main areas of insight we have are feeding and reproduction.
Pterosaur anatomy give several clues as how they may have fed, but inevitably such interpretations remain tentative and often controversial.

Rynchops skimming. While it is possible some smaller pterosaurs fed like this, it is now thought unlikely for large ones.
Many pterosaur appear to have eaten fish—through their exact methods are not known, and probably varied from species to species. Skimming, in the manner of Rynchops (skimmers) has been suggested for many pterosaurs, but recent studies have found that this is an unlikely method of feeding for large pterosaurs (Humphries et al 2007). Dive-fishing is also possible for small pterosaurs—but again, unlikely for large ones, as it seems doubtful that they could take off from the water. So it seems the most like fishing method for medium to large pterosaurs was to snatch fish on the wing, with minimal contact with the water.
Other pterosaurs show specialised adaptations for other types of feeding. For example, agnuronathids had wide, frog-like jaws, and combined with their small size and habitat, suggests they probably fed on flying insects. Pterodaustro had a long, up-curved snout with thousands of bristle-like teeth, which it probably used to sieve small organisms out of the water, much like a flamingo. Azhdarchids may have been terrestially adapted, and foraged for small animals and carrion (Witton & Naish 2008).
Analysis of pterosaur eggs suggests they buried them like crocodiles and turtles. Pterosaur hatchlings were reasonably well-developed, suggesting they were able to fly from a young age. This suggests that pterosaurs did not practice as much parental care as birds, and may not have cared for their young at all (Unwin 2003).
Restoring Palaeo-environments
Most pterosaurs are known from shoreline or marine deposits, meaning they lived near oceans or inland seas. However, many pterosaurs are also known from terrestrial deposits.
It can be difficult to get information on the plant-life contemporary with pterosaurs. Generally it is necessary to research the primary literature, as overviews are rare (and often non-existent). Unfortunately, the literature is often not illustrated in a manner helpful to an artist. Comparisons with the closest living relatives is invaluable.