From belly-dragging sprawlers to two-legged dynamos and everything in-between: terrestrial locomotion in pterosaurs
By Mark Witton
It didn’t take long for early pterosaur workers to figure out that pterosaurs were flying animals. Their hyperelongate forelimbs, fused torso elements, robust shoulder girdle and pectoral elements are clear indications that pterosaurs were adapted for self-propelled flight and, although some arguments have raged over the finer details of their flight styles, no-one has questioned their basic volant ability. However, the same cannot be said for pterosaur terrestrial ability: for almost as long as pterosaurs have been known, little consensus has existed on both how and how well pterosaurs would be able to move on land. Early workers thought pterosaurs were probably relatively confident terrestrial locomotors, although no-one could agree whether they would stand in a bird-like, bipedal fashion, a lizard-like quadrupedal configuration or an erect, mammal-like quadrupedal stance. Pterosaurologists of the mid-20th century took a dimmer view: these workers suggested that pterosaurs could barely stand, let alone walk, and would have to push themselves over the ground with dragging bellies and useless forelimbs held aloft (Fig 1A).
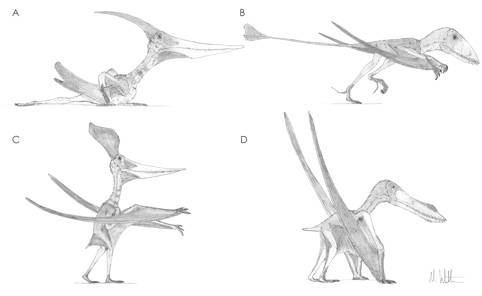
Fig. 1. Theories of terrestrial locomotion in pterosaurs. A, the belly-dragging Pteranodon of championed in the 1970s; B, a bipedal Dimorphodon of the early 1980s; C, semi-erect Pteranodon proposed in 1990; D, a sprawling Anhanguera the late 1980s and early 1990s. Note that the wing membrane configurations reflect the views of the respective authors at the time of their publications and not current thinking of pterosaur wing membrane distributions.
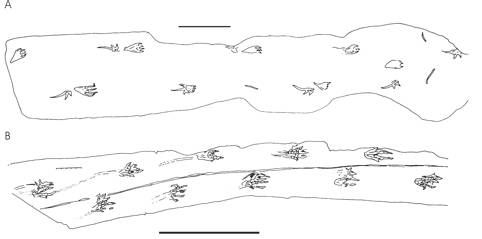
Fig. 2. Pteraichnus and Caiman tracks. A, Pteraichnus saltwashensis; B, trackway produced by a running Caiman sclerops. Note the distinctions in print morphology, tail-drags and print placement. Scale bars represent 200 mm.
Opinions changed in the 1980s when several pterosaur workers attempted biomechanical studies of pterosaur limbs in efforts to figure out their terrestrial competence: alas, this confused matters further as different interpretations of the same material led to drastically different conclusions. In the space of a decade, pterosaurs were restored as dinosaur-like bipeds (Fig. 1B), penguin-like bipeds (Fig. 1C), sprawled-limbed quadrupeds (Fig. 1D) and erect-limbed quadrupeds. Ironically, workers at this time were ignoring the Rosetta Stone of this mystery, a discovery that had been made several decades before but largely ignored: pterosaur trackways (Fig. 2A). These distinctive tracks, named Pteraichnus, were dismissed as being made by crocodilians (on rather shaky evidence – compare Fig. 2A with 2B and judge for yourself) and were not used by pterosaur workers to assess pterosaur terrestrial locomotion until comparatively recently. In fact, it wasn’t until the mid-1990s that the pterosaur origin of Pteraichnus was firmly re-established and, along with the discovery of other pterosaur tracks across the world (including the Cretaceous ichnogenera Purbeckopus from Southern Britain and Haenamichnus from Southern Korea; Fig. 3) a picture of how pterosaurs, big and small, stood, walked and ran could be painted.
First and foremost, Pteraichnus and its ichno-kin demonstrate to us that pterosaurs walked quadrupedally on digitigrade forelimbs and plantigrade hindlimbs. Only the first three fingers regularly touch the floor as the flight finger was apparently folded and stowed alongside the body when grounded, occasionally leaving a fourth digit trace in the manus print. Folding the wing in this way requires the metacarpals to be medially rotated at the wrist, thereby forcing the manual digits to produce laterally- and posterolaterally directed digit impressions. The comparatively shallow pes prints have a narrow ankle that, in Haenamichnus at least, appears to have borne a large, fleshy pad with toes that often curve medially due to the presence of long toe claws.
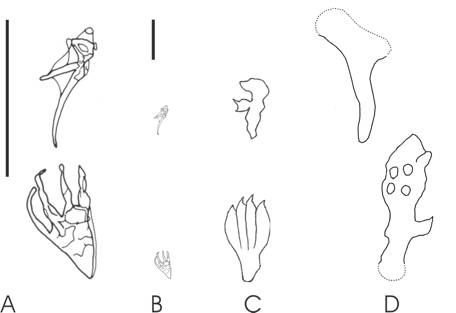
Fig. 3. Pterosaur manus (top row) and pes (bottom) prints. A, Pteraichnus saltwasensis; B, P. saltwashensis in scale with C and D; C, Purbeckopus pentadactylus, D, Haenamichnus uhangriensis. Scale bars represent 100 mm.
Now, the position of these prints relative to one another is perhaps the most interesting aspect of pterosaur trackways. It was once though that the disproportionately long forelimbs of pterosaurs should lead to them having a splayed forelimb configuration when standing and walking, but pterosaur manus prints fall almost directly anterior to the pes prints with only slightly more pronounced displacement from the midline. Hence, reconstructing a pterosaur standing on such trackways shows that both the hindlimb and forelimb are not splayed anywhere near as much as suggested by earlier workers. In fact, the limbs of pterosaurs are held parasagitally (Fig. 4), a feat achieved by retracting the humerus posteriorly and ventrally in the glenoid and the highly mobile joints of the acetabulum and knee joints. This gives pterosaurs a relatively efficient stance with their bodyweight carried on near-columnar limbs rather than the energy-sapping stance of sprawling reptiles and amphibians. However, some trackways show that pterosaurs did sometimes splay their forelimbs, a track feature that correlates well with increased stride length. It therefore seems that pterosaurs splayed their forelimbs when walking quickly or running, a consequence of rotating the body forward to give extra reach to the shoulders. In such a posture, the forelimbs achieved greater reach but were not given sufficient ground-clearance to reach the midline. Conversely, pterosaurs moving at more sedate paces would have a more erect stance that allowed the forelimbs to be brought in closer to the midline at the consequence of a comparatively limited reach.
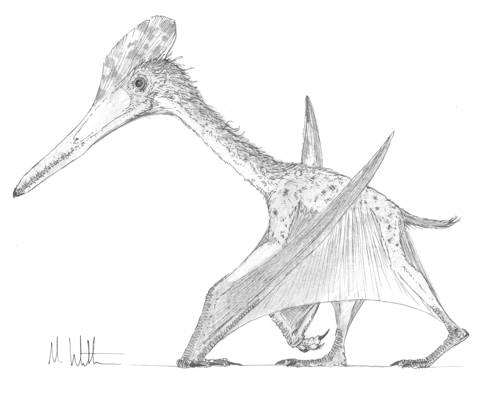
Fig. 4. Pterodactylus demonstrating the erect, parasagittal gait of grounded pterodactyloids.
Additional evidence that pterosaurs could move around competently without flight is the discovery of traces left by a swimming pterosaur (partial pes prints and a plethora of scratch marks made by pterosaurs kicking their way through shallow water). Wading behaviour may also be recorded: bizarre pterosaur trackways comprised of manus prints alone have been suggested to be created by pterosaurs wading in shallow water with buoyed-up hindquarters. An alternative explanation for these strange tracks is the front-heavy structure of standing pterosaurs: being flying animals, pterosaurs were weightier up-front with their wings and flight musculature anchored around their shoulder regions. Hence, their centre of gravity is shifted forward and puts more weight on their forelimbs than hindlimbs, thereby creating ‘manus-only’ trackways on substrata that only yields to the relatively heavy footfall of the forelimb. Whatever the explanation for these tracks, it’s quite clear that pterosaurs were highly-competent terrestrial animals, able to explore all the nooks and crannies of Mesozoic landscapes without having to resort to flight. However, there is reason to believe that this is not the case for all pterosaurs: there are several reasons to believe that pterodactyloids and basal pterosaurs had drastically different postures and terrestrial capabilities.
Y’see, pterosaur tracks are only found in strata younger than the middle Jurassic, but pterosaurs have a body fossil record extending well into the upper Triassic. Why, then, do we have millions of years of Triassic and Jurassic pterosaur body fossils with no corresponding ichnological record? The answer may lie in the broad anatomical distinction between basal pterosaurs and pterodactyloids: exceptionally well preserved basal pterosaurs appear to have a single, broad cruropatagia that extends fully between the hindlimbs and pelvis with its ventral margin supported by the elongated fifth toes of the pedes. Pterodactyloids, by contrast, appear to have two reduced, independent cruopatagia that run along the posterior margin of the legs, thereby liberating the left and right pedes from each other. Hence, basal pterosaurs essentially had to run around with their shoelaces tied together while pterodactyloids enjoyed uninhibited, independent use of each leg. Given this, it is perhaps unsurprising that the start of the pterosaur track record coincides with the evolution of pterodactyloids themselves. Their relatively limited terrestrial abilities may have persuaded basal pterosaurs to avoid wandering around on the ground if they could, thereby providing little opportunity to create trackways.
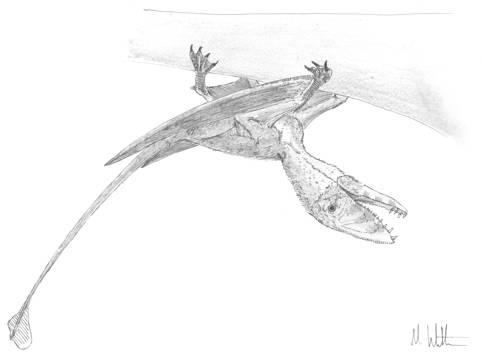
Fig. 5. Dimorphodon climbs. Note the developed foot and hand claws, the sprawling hindlimbs and low centre of gravity.
Now, this is not to say that basal pterosaurs were locomotory inept from the moment they landed. They may, however, have spent more time running around trees and cliffs than over floodplains and tidal flats. Basal pterosaurs typically have deepened, highly recurved manual and pedal claws with comparatively large flexor tubercles compared to the relatively slender claws of pterodactyloids. These claws are extremely thin despite their depth and would make excellent crampons to provide purchase when climbing, especially when combined with the antungual sesamoids and elongate penultimate phalanges that characterise the hands and feet of many basal forms. Furthermore, the orientation of the femoral head in basal pterosaurs means that the femur is projected forward, upward and laterally from the acetabulum, thereby causing the sprawling gait for the hindlimbs that acted in concert with the relatively short metacarpals to bring the bodies of these pterosaurs close to any surface they happened to be climbing over. These are all excellent adaptations to climbing (Fig. 5), and we should expect early Mesozoic environments to be covered with pterosaurs hanging from cliff faces, tree trunks and branches. What’s more, the restriction on independent hindlimb movement created by the broad curuopatagium does not necessarily mean that basal pterosaurs were slow movers: by moving their hindlimbs together they may have saltated along like leathery-winged, bucked-toothed squirrels. Basal pterosaurs could probably out-climb their pterodactyloid descendents, in fact: while some pterodactyloids possess some climbing characteristics, their feet and hands are relatively underdeveloped compared to those of basal pterosaurs and their longer distal limb elements would make them less stable climbers. Hence, while pterodactyloids dominated pterosaur ecomorphospace on horizontal surfaces, basal pterosaurs ruled the vertical.
So there you go, then: pterosaur terrestrial locomotion in a nutshell. Of course, while our understanding of this aspect of pterosaur palaeobiology has improved considerably in the last decade or so, many questions remain unanswered. How, for instance, did the fingerless (save for the flight digit) Nyctosaurus locomote when grounded, and just how did those crazy euornithocheiroids compensate for their grossly disproportionate forelimb-hindlimb ratios? When, why and how did basal pterosaurs stop hopping around trees and cliffs to become terrestrialised pterodactyloids? And will we ever, ever find a basal pterosaur trackway? Although the answers to all of these are frustratingly obscured at present, we can at least be pretty confident that after centuries of controversy, we’ve finally got a picture of pterosaur terrestrial locomotion that brings all lines of evidence together. We are, then, setting off on the right foot to answer the reams of questions that lie in the finer details.